Understanding Peptide Half-Life: t1/2, Steady State and Accumulation
Dr. Sieglinde Klaus
Scientific Editorial Team · Bergdorf Bioscience

Dr. Sieglinde Klaus
Scientific Editorial Team · Bergdorf Bioscience
The half-life (t1/2) of a peptide describes the time it takes for the plasma concentration to fall to half of its starting value. Under the usual first-order kinetics this value is concentration-independent and governs how fast a substance is eliminated, when a steady state is reached, and how strongly repeated doses add up. This article explains the concepts in purely scientific terms, strictly for research purposes.
The elimination half-life is defined as the time required for the concentration of a substance in the body to drop to half of its initial value (Hallare & Gerriets, 2025). For a peptide with a t1/2 of 24 hours, 50 percent of the starting amount would still be measurable after one day, 25 percent after two days, and 12.5 percent after three. This halving decline is the hallmark of first-order kinetics, where the amount eliminated per unit time is proportional to the concentration currently present.
Peptides differ enormously. The gastric pentadecapeptide BPC-157 showed an elimination half-life of only about 15.2 minutes in rats after intravenous administration of 20 micrograms per kilogram (He et al., 2022). Modified peptides such as semaglutide, by contrast, reach around 160 hours, roughly one week (Yang et al., 2024). This span from minutes to weeks is why half-life is the central parameter of any pharmacokinetic analysis. You can visualise the actual time course for a selection of research peptides with our half-life calculator.
Most clinically relevant substances, including most peptides in the physiological concentration range, follow first-order kinetics. Here a constant percentage of the amount present is eliminated per unit time, not a constant absolute amount. The concentration therefore falls exponentially: C(t) = C0 times e to the power of minus k times t, where k is the elimination rate constant. The relationship between k and half-life is t1/2 = 0.693 divided by k, because ln(2) is roughly 0.693.
The practical value of this relationship is that knowing the half-life means knowing the entire decay curve. A peptide with a t1/2 of 12 hours has an elimination rate constant of about 0.0578 per hour, so roughly 5.6 percent of whatever amount remains disappears each hour. The constant k can be obtained in reverse from the half-life by dividing 0.693 by t1/2, so the two quantities are always interconvertible and describe exactly the same elimination dynamics.
The exponential character becomes visually clear when concentration is plotted on a logarithmic scale: the curved decay line then becomes a straight line whose slope equals the negative rate constant. This linearisation is why pharmacokineticists often display elimination data semilogarithmically, since the half-life can be read directly from the slope. Important is the distinction from zero-order kinetics, where a fixed absolute amount is eliminated regardless of concentration (the classic example is ethanol). Under that kinetics the concentration falls not exponentially but linearly, and the notion of half-life loses its constancy, because the apparent half-life then depends on the starting concentration. Such saturation effects only appear when the eliminating enzymes or transporters become overloaded. For the dose ranges common in research, the first-order assumption usually holds and forms the basis of all the calculation models used in this article.
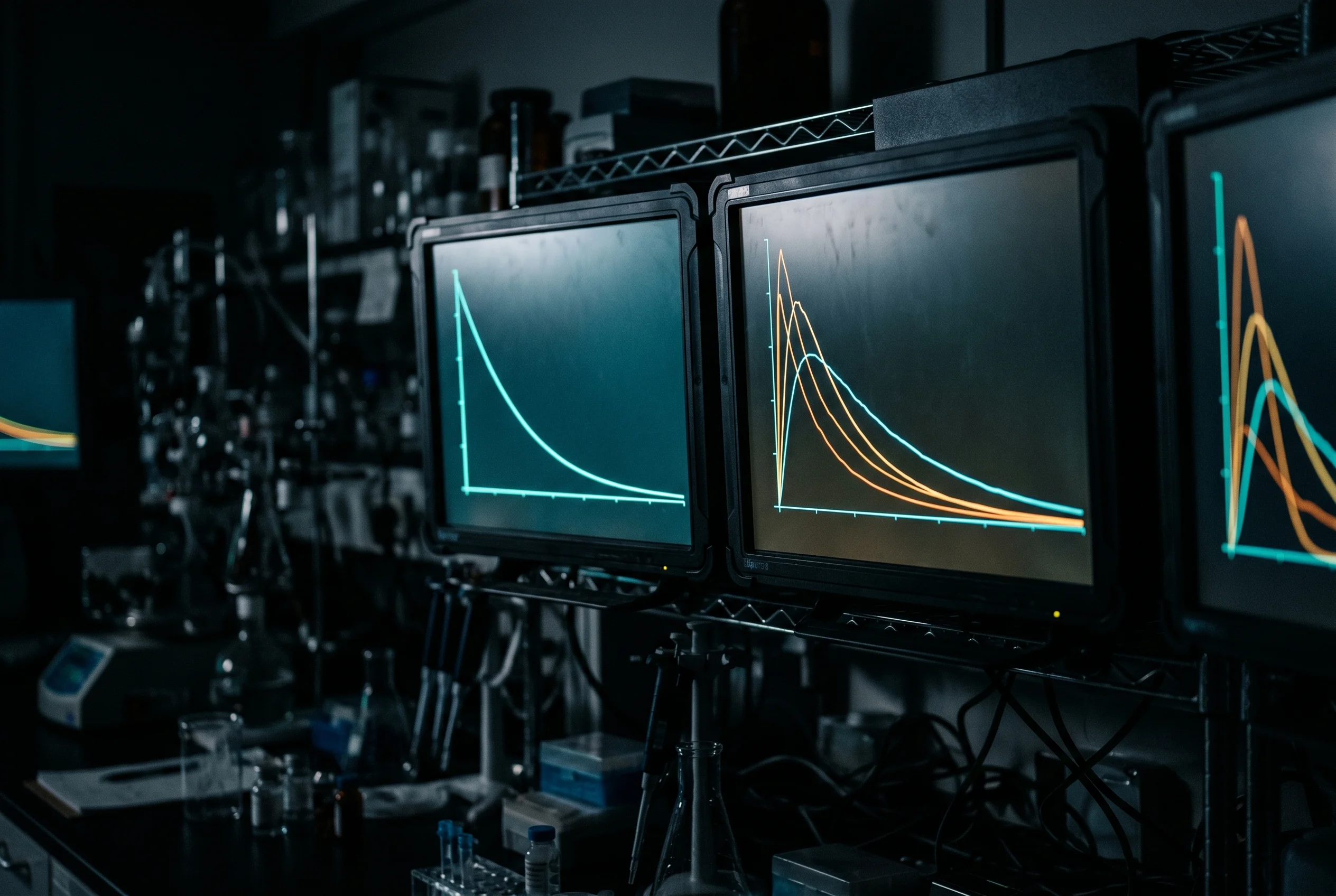
Because the decline is exponential, the concentration mathematically never reaches exactly zero, but it falls below a practically meaningful threshold very quickly. The common rule of thumb is that after four to five half-lives a substance is considered effectively eliminated, since only about 3 to 6 percent of the starting amount remains (Hallare & Gerriets, 2025).
The numbers in detail: after one half-life, 50 percent remains; after two, 25 percent; after three, 12.5 percent; after four, 6.25 percent; and after five, 3.125 percent. Applied to real peptides this means very different time windows. Tirzepatide, with a half-life of about five days (Schneck et al., 2024), would be largely gone from the system after roughly 20 to 25 days. BPC-157, with its roughly 15 minutes, would be practically undetectable after a little over an hour. This range makes clear that statements about residence time must always be made relative to the specific half-life. Blanket time figures without reference to the actual molecule are not scientifically defensible.
A common misconception is equating plasma half-life with duration of effect. The plasma half-life describes only how quickly the measurable concentration in the blood declines. The functional or pharmacodynamic half-life, by contrast, describes how long a measurable biological effect persists. The two can diverge considerably when a peptide binds to tissue receptors, is released there with delay, or triggers a signalling cascade that lasts longer than the substance itself is detectable.
The reason lies in the spatial separation of the compartments: what is measured in plasma is only the freely circulating fraction. Part of the peptide moves into the so-called deep compartment, meaning poorly perfused tissues or receptor-bound reservoirs, from which it returns only slowly. As long as this back-flow continues, a biological effect persists even though the plasma concentration may already have fallen below the limit of detection. The functional half-life is therefore in practice often longer than the plasma half-life, and exactly this difference explains why some peptides show measurable effects for hours or days despite a short plasma residence. For precise research models it is therefore always necessary to state whether plasma or functional kinetics is being discussed, because the rules of thumb for elimination strictly refer only to the plasma half-life.

The enormous spread of half-lives, from minutes for native peptides to a week for modern agents, is no accident but the result of deliberate molecular modifications. Native peptides are rapidly cleaved by peptidases in the body and filtered by the kidneys, because their molecular mass usually lies below the glomerular filtration threshold. Both routes can be slowed dramatically by attaching a fatty-acid chain, the process known as acylation or lipidation (Menacho-Melgar et al., 2018).
The principle rests on reversible binding to albumin, the most abundant plasma protein. A fatty acid coupled to the peptide inserts itself into the fatty-acid binding pockets of albumin. The bound peptide is thereby too large for renal filtration and is sterically shielded from enzymatic degradation; albumin acts as a circulating depot from which the free, active form is slowly released (Menacho-Melgar et al., 2018). Through this principle semaglutide is more than 99 percent bound to albumin and carries a C18 fatty-diacid side chain that raises the half-life to around 160 hours (Yang et al., 2024). Tirzepatide is about 80 percent bound to plasma proteins and distributes in a volume of distribution of roughly 10.3 litres (Schneck et al., 2024). These mechanisms explain why the half-life measured in blood is inseparably tied to the chemical structure of the molecule and cannot be transferred from one peptide to another. Even the substitution of single amino acids that renders a molecule resistant to cleavage by dipeptidyl peptidase-4 can extend the residence time many times over.
When repeated doses are given at regular intervals, each new amount adds to whatever remains in the system from previous doses. As long as more is added than eliminated in each interval, the mean concentration rises. Eventually input and elimination balance out: a dynamic equilibrium establishes, the so-called steady state. A pharmacological rule of thumb states that this state is reached after about five half-lives (Wadhwa & Cascella, 2023).
This rule of thumb follows directly from the exponential function: with each further half-life the mean concentration closes half of the remaining gap to the plateau. After one half-life about 50 percent of the plateau is reached, after two about 75 percent, after three about 87.5 percent, and after five already more than 96 percent. The very same pattern that describes the decay of a single dose governs, in mirror image, the build-up to steady state, which is why both processes take exactly equally long.
Key is a frequently misunderstood property: the time to steady state depends solely on the half-life, not on the dose level. A higher dose leads to a higher plateau but not to faster attainment of equilibrium. For tirzepatide, with about five days half-life, steady state is therefore only reached after roughly four weeks of weekly dosing (Schneck et al., 2024). For peptides with a very short half-life of a few minutes, by contrast, near-complete elimination occurs after each single dose, so hardly any classic plateau builds up. The concept of steady state is thus only relevant for substances whose half-life is in the range of the dosing interval or longer. Anyone wishing to reach the plateau faster without permanently raising the maintenance dose would have to use a single loading dose, which is, however, a separate concept beyond the pure half-life analysis.
The accumulation factor (Rac) quantifies how much the steady-state concentration rises relative to a single dose. The basic formula is Rac = 1 divided by (1 minus the fraction remaining in the interval), or equivalently 1 divided by the fraction eliminated in the interval. Expressed through the half-life it becomes Rac = 1 divided by (1 minus 0.5 to the power of (dosing interval divided by t1/2)). Equivalently, the fraction remaining in the interval follows from e to the power of minus k times tau, where tau is the dosing interval.
A concrete example illustrates this. If the dosing interval equals exactly one half-life, 50 percent remains at the end of each interval. The accumulation factor is then 1 divided by (1 minus 0.5), which equals 2, so the steady-state concentration is about twice that of a single dose. If the interval is twice the half-life, 25 percent remains and Rac is roughly 1.33. For very short intervals relative to the half-life the factor rises sharply; with an interval of a quarter of the half-life, for instance, to about 6.3. Real data fit this model: tirzepatide showed a mean accumulation of about 1.7-fold with weekly dosing, consistent with a half-life of around five days and a seven-day interval (Schneck et al., 2024). The longer the half-life relative to the interval, the stronger the accumulation.
In pharmacokinetic practice the half-life is the most important guide for choosing the spacing between repeated doses in a research protocol. An interval much shorter than the half-life leads to strong accumulation and high plateau concentrations. An interval that is a multiple of the half-life lets the concentration fall far between doses and creates large swings between peak and trough.
The reason for this relationship is purely mathematical: the ratio of peak to trough concentration at steady state is determined solely by the ratio of dosing interval to half-life. If the interval equals one half-life, the concentration halves between two doses, so the peak-to-trough ratio is about 2 to 1. With an interval of four half-lives the concentration falls to one sixteenth, so the fluctuation becomes extreme. The long half-lives of modern peptides in particular therefore explain their dosing schemes. Semaglutide, with about one week half-life (Yang et al., 2024), and tirzepatide, with about five days (Schneck et al., 2024), allow weekly intervals because the concentration does not fall too far between two doses. Short-lived peptides such as BPC-157, with a minute-scale half-life (He et al., 2022), would practically vanish between doses on the same schedule. The half-life therefore sets the frame within which a sensible ratio of fluctuation and accumulation is possible. Researchers working with several peptides can model amounts with the peptide calculator and time courses with the half-life calculator.
A fully worked example ties the preceding concepts together. Suppose a research peptide has a half-life of 48 hours and is given in the same amount every 24 hours. First the elimination rate constant: k = 0.693 divided by 48, which is roughly 0.0144 per hour. The fraction remaining in the 24-hour interval is e to the power of minus 0.0144 times 24, about 0.707, so roughly 70.7 percent remains.
From this follows the accumulation factor: Rac = 1 divided by (1 minus 0.707), about 3.4. The steady-state concentration is therefore around 3.4 times what a single dose produces. Until this steady state is reached, roughly five half-lives pass, about 240 hours or ten days. Extend the interval to 48 hours, exactly one half-life, and the fraction remaining drops to 50 percent while Rac falls to 2. Shorten it instead to 12 hours, a quarter of the half-life, and the fraction remaining rises to about 0.841 while Rac climbs to roughly 6.3. This numerical exercise vividly shows how sensitively accumulation responds to the ratio of interval to half-life. The half-life calculator performs exactly these calculations and plots the curve, steady state and accumulation factor graphically. For a foundational introduction to this class of molecules, see the article What are peptides?.
All the calculations so far rest on the one-compartment model, which treats the body as a single, homogeneously mixed space from which the substance disappears with a single rate constant. This model is elegant because it gets by with a single half-life, and it is accurate enough for many research purposes. It captures reality only approximately, however, because it assumes that a peptide distributes instantly and uniformly throughout the entire volume of distribution immediately after dosing.
In fact many peptides do not distribute instantaneously. They first flow rapidly into well-perfused tissues and only slowly into poorly perfused compartments. This produces a two-phase course: a steep distribution phase shortly after the dose, followed by a flatter elimination phase in which the concentration is maintained by redistribution from the tissues. Such behaviour is described more accurately by the two-compartment model, which then yields two different half-lives, a short distribution half-life and a longer terminal half-life. Anyone considering only a single number easily confuses these two phases and underestimates the residence time, because the terminal phase governs tissue residence, whereas the distribution phase reflects only the initial, rapid drop in plasma concentration.
This is precisely where the practical consequence for modelling lies: the four-to-five-half-lives rule refers to the terminal half-life, not the fast distribution phase. Anyone who wrongly inserts the distribution half-life would dramatically underestimate the elimination time. Further limits concern nonlinear kinetics at saturation of the eliminating systems, active metabolites that are themselves effective and extend the effective duration, and pronounced tissue binding that lengthens the terminal phase. The assumption of instantaneous, complete absorption used in the worked examples also holds only for intravenous administration; with subcutaneous delivery the absorption delays the course further and can lengthen the apparent half-life, an effect known as flip-flop kinetics. For most research peptides in the usual dose range the one-compartment model nonetheless remains a usable approximation, but one should remain aware of the simplification and not overstretch the rules of thumb.
Not necessarily. A long half-life smooths concentration fluctuations and allows less frequent dosing, but it also lengthens the time to steady state and the residence time after discontinuation. Which property is desirable depends entirely on the research objective and cannot be answered in blanket terms.
Under first-order kinetics the half-life is concentration-independent and stays constant across the usual dose range. Only when the eliminating systems saturate, that is, when kinetics shift to zero order, can the apparent half-life become dose-dependent. For most research peptides this is not relevant.
Targeted molecular modifications such as amino-acid substitutions or the attachment of fatty-acid chains increase albumin binding and protect against enzymatic degradation (Menacho-Melgar et al., 2018). Semaglutide, for instance, carries a C18 fatty-diacid side chain that extends the half-life to around 160 hours (Yang et al., 2024).
It is a good approximation for substances with first-order kinetics but has exceptions (Wadhwa & Cascella, 2023). Substances with nonlinear kinetics, pronounced tissue binding or active metabolites can deviate markedly. The rule therefore does not replace substance-specific modelling.
For research purposes only. Not intended for human consumption. Scientific editor: Dr. Sieglinde Klaus
