Peptide-halfwaardetijd begrijpen: t1/2, steady state en accumulatie
Dr. Sieglinde Klaus
Wetenschappelijke redactie · Bergdorf Bioscience

Dr. Sieglinde Klaus
Wetenschappelijke redactie · Bergdorf Bioscience
De halfwaardetijd (t1/2) van een peptide beschrijft de tijd waarin de plasmaconcentratie tot de helft van de uitgangswaarde daalt. Bij de gebruikelijke eersteordekinetiek is deze waarde concentratieonafhankelijk en bepaalt zij hoe snel een stof wordt geelimineerd, wanneer zich een steady state instelt en hoe sterk herhaalde toedieningen zich opstapelen. Dit artikel verklaart de concepten zuiver wetenschappelijk, uitsluitend voor onderzoeksdoeleinden.
De eliminatiehalfwaardetijd is gedefinieerd als de tijdspanne waarin de concentratie van een stof in het lichaam tot de helft van de uitgangswaarde daalt (Hallare & Gerriets, 2025). Bij een peptide met een t1/2 van 24 uur zou na een dag nog 50 procent van de oorspronkelijke hoeveelheid meetbaar zijn, na twee dagen 25 procent en na drie dagen 12,5 procent. Deze halverende afname is het kenmerk van eersteordekinetiek, waarbij de per tijdseenheid geelimineerde hoeveelheid evenredig is met de op dat moment aanwezige concentratie.
Peptiden verschillen enorm: het gastrische pentadecapeptide BPC-157 vertoonde bij ratten na intraveneuze toediening van 20 microgram per kilogram een eliminatiehalfwaardetijd van slechts ongeveer 15,2 minuten (He et al., 2022). Gemodificeerde peptiden zoals semaglutide bereiken daarentegen rond de 160 uur, dus ongeveer een week (Yang et al., 2024). Deze spreiding van minuten tot weken is de reden waarom de halfwaardetijd de centrale parameter is van elke farmacokinetische beschouwing. Het concrete verloop kunt u met onze halfwaardetijd-calculator voor een selectie van onderzoekspeptiden visualiseren.
De meeste klinisch relevante stoffen, waaronder de meeste peptiden in het fysiologische concentratiebereik, volgen een eersteordekinetiek. Daarbij wordt steeds een constant percentage van de aanwezige hoeveelheid per tijdseenheid geelimineerd, niet een constante absolute hoeveelheid. De concentratie daalt daarom exponentieel: C(t) = C0 maal e tot de macht min k maal t, waarbij k de eliminatiesnelheidsconstante is. Tussen k en de halfwaardetijd geldt het verband t1/2 = 0,693 gedeeld door k, omdat ln(2) ongeveer 0,693 bedraagt.
Het praktische nut van dit verband: kent men de halfwaardetijd, dan kent men het volledige uitdovingsverloop. Een peptide met een t1/2 van 12 uur heeft een eliminatiesnelheidsconstante van ongeveer 0,0578 per uur. Per uur verdwijnt dus rond de 5,6 procent van de telkens nog aanwezige hoeveelheid. Wiskundig kan de constante k omgekeerd uit de halfwaardetijd worden afgeleid door 0,693 door t1/2 te delen, zodat beide grootheden op elk moment in elkaar omrekenbaar zijn en steeds dezelfde eliminatiedynamiek beschrijven.
Het exponentiele karakter wordt aanschouwelijk wanneer men de concentratie logaritmisch uitzet: uit de gekromde uitdovingskromme ontstaat dan een rechte lijn waarvan de helling overeenkomt met de negatieve snelheidsconstante. Deze linearisatie is de reden waarom farmacokinetici eliminatiegegevens vaak halflogaritmisch weergeven, want uit de helling kan de halfwaardetijd direct worden afgelezen. Belangrijk is het onderscheid met de nulde-ordekinetiek, waarbij onafhankelijk van de concentratie een vaste absolute hoeveelheid wordt geelimineerd (klassiek voorbeeld: ethanol). Bij die kinetiek daalt de concentratie niet exponentieel, maar lineair, en verliest het begrip halfwaardetijd zijn constantheid, omdat de schijnbare halfwaardetijd dan van de uitgangsconcentratie afhangt. Dergelijke verzadigingseffecten treden pas op wanneer eliminerende enzymen of transporters zijn uitgeput. Voor de in onderzoek gebruikelijke doseringen is de aanname van de eerste orde in de regel houdbaar en vormt zij de grondslag van alle rekenmodellen die in dit artikel worden gebruikt.
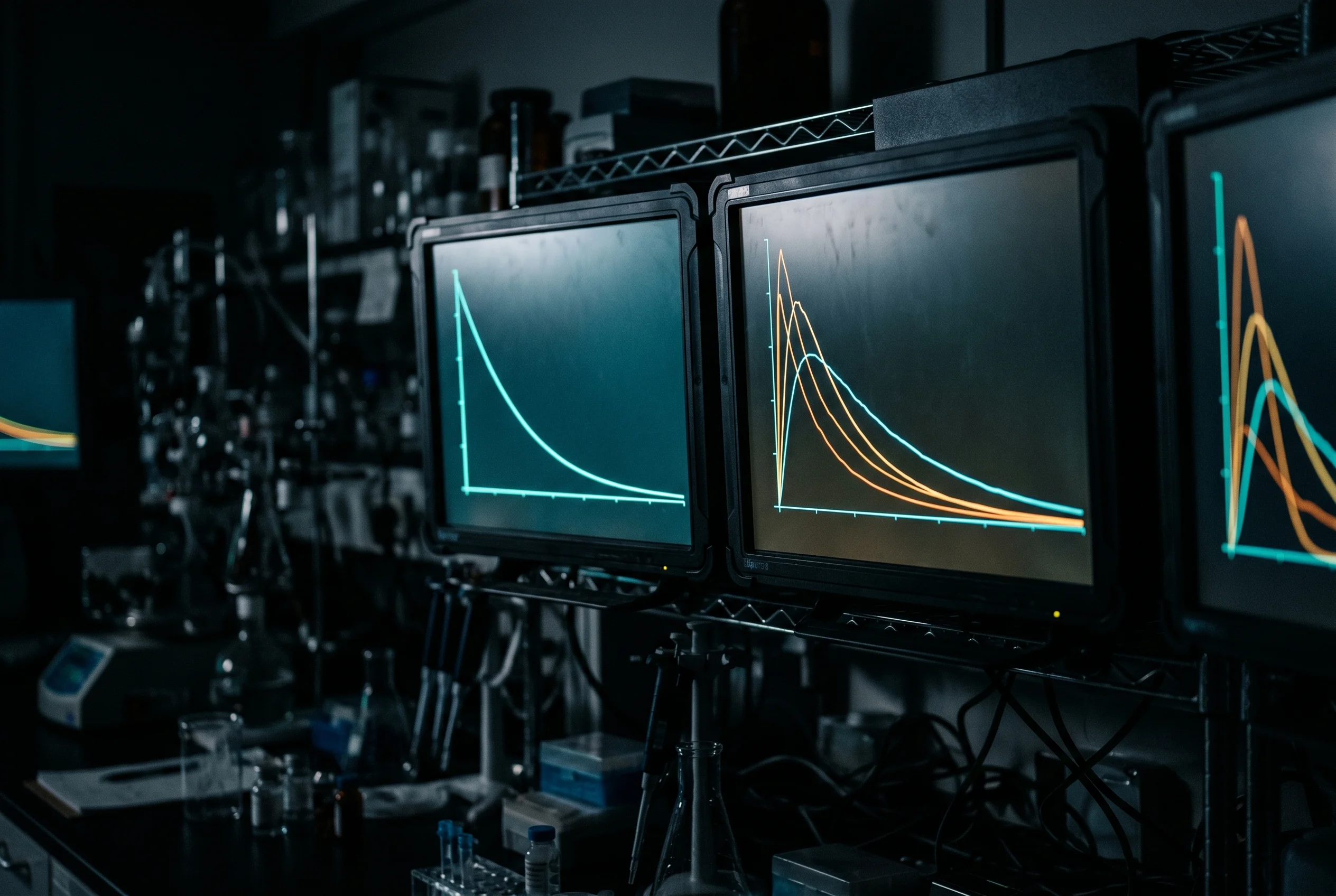
Omdat de afname exponentieel verloopt, bereikt de concentratie wiskundig nooit exact nul, maar daalt zij zeer snel onder een praktisch betekenisvolle drempel. De gangbare vuistregel luidt: na vier tot vijf halfwaardetijden geldt een stof als effectief geelimineerd, omdat er dan nog maar ongeveer 3 tot 6 procent van de oorspronkelijke hoeveelheid aanwezig is (Hallare & Gerriets, 2025).
De cijfers in detail: na een halfwaardetijd resteert 50 procent, na twee 25 procent, na drie 12,5 procent, na vier 6,25 procent en na vijf 3,125 procent. Toegepast op echte peptiden betekent dit zeer uiteenlopende tijdvensters. Tirzepatide met een halfwaardetijd van ongeveer vijf dagen (Schneck et al., 2024) zou na rond de 20 tot 25 dagen grotendeels uit het systeem verdwenen zijn. BPC-157 met zijn ongeveer 15 minuten zou daarentegen na ruim een uur praktisch niet meer aantoonbaar zijn. Deze spanwijdte maakt duidelijk dat uitspraken over verblijfsduur altijd relatief aan de betreffende halfwaardetijd moeten worden gedaan. Algemene tijdsaanduidingen zonder verwijzing naar het concrete molecuul zijn wetenschappelijk niet houdbaar.
Een veelvoorkomend misverstand is het gelijkstellen van plasmahalfwaardetijd en werkingsduur. De plasmahalfwaardetijd beschrijft uitsluitend hoe snel de meetbare concentratie in het bloed afneemt. De functionele of farmacodynamische halfwaardetijd beschrijft daarentegen hoe lang een meetbaar biologisch effect aanhoudt. Beide kunnen aanzienlijk uiteenlopen wanneer een peptide aan weefselreceptoren bindt, daar vertraagd wordt vrijgesteld of een signaalcascade in gang zet die langer aanhoudt dan de stof zelf aantoonbaar is.
De reden ligt in de ruimtelijke scheiding van de compartimenten: wat in het plasma wordt gemeten, is slechts het vrij circulerende aandeel. Een deel van het peptide trekt het zogenaamde diepe compartiment in, dus slecht doorbloede weefsels of receptorgebonden reservoirs, waaruit het slechts langzaam terugstroomt. Zolang deze terugstroom aanhoudt, blijft er een biologische werking bestaan, hoewel de plasmaconcentratie reeds onder de detectiegrens kan zijn gedaald. De functionele halfwaardetijd is daarom in de praktijk vaak langer dan de plasmahalfwaardetijd, en juist dit verschil verklaart waarom sommige peptiden ondanks een korte plasmaverblijfsduur urenlang of dagenlang meetbare effecten vertonen. Voor nauwkeurige onderzoeksmodellen moet daarom steeds worden aangegeven of er over plasma- of functionele kinetiek wordt gesproken, want de vuistregels voor eliminatie hebben strikt genomen alleen betrekking op de plasmahalfwaardetijd.

De enorme spanwijdte van de halfwaardetijden, van minuten bij natieve peptiden tot een week bij moderne werkzame stoffen, is geen toeval, maar het resultaat van gerichte moleculaire modificaties. Natieve peptiden worden in het lichaam snel door peptidasen gesplitst en via de nieren gefiltreerd, omdat hun molecuulmassa meestal onder de glomerulaire filtratiedrempel ligt. Beide routes laten zich door het aanhechten van een vetzuurketen, de zogenaamde acylering of lipidering, drastisch vertragen (Menacho-Melgar et al., 2018).
Het principe berust op de omkeerbare binding aan albumine, het meest voorkomende plasma-eiwit. Een aan het peptide gekoppeld vetzuur nestelt zich in de vetzuur-bindingsplaatsen van het albumine. Het gebonden peptide is daardoor te groot voor de renale filtratie en sterisch beschermd tegen de enzymatische afbraak; albumine werkt als een circulerend depot waaruit de vrije, werkzame vorm langzaam wordt vrijgesteld (Menacho-Melgar et al., 2018). Semaglutide is via dit principe voor meer dan 99 procent aan albumine gebonden en draagt een C18-vetzuurdizuur-zijketen die de halfwaardetijd tot rond de 160 uur verhoogt (Yang et al., 2024). Tirzepatide is voor ongeveer 80 procent aan plasma-eiwitten gebonden en verdeelt zich over een verdelingsvolume van ongeveer 10,3 liter (Schneck et al., 2024). Deze mechanismen verklaren waarom de in het bloed gemeten halfwaardetijd onlosmakelijk samenhangt met de chemische structuur van het molecuul en zich niet van het ene peptide naar het andere laat overdragen. Reeds de vervanging van afzonderlijke aminozuren, die een molecuul ongevoelig maakt voor de splitsing door dipeptidylpeptidase-4, kan de verblijfsduur vele malen verlengen.
Worden herhaalde toedieningen met regelmatige tussenpozen verstrekt, dan telt elke nieuwe hoeveelheid op bij wat er van eerdere toedieningen nog in het systeem zit. Zolang er in elk interval meer wordt toegevoerd dan geelimineerd, stijgt de gemiddelde concentratie. Op een gegeven moment houden toevoer en eliminatie elkaar in evenwicht: er stelt zich een dynamisch evenwicht in, de zogenaamde steady state. Een farmacologische vuistregel zegt dat deze toestand na ongeveer vijf halfwaardetijden wordt bereikt (Wadhwa & Cascella, 2023).
Deze vuistregel volgt rechtstreeks uit de exponentiele functie: na elke verdere halfwaardetijd nadert de gemiddelde concentratie het plateau met de helft van de nog resterende afstand. Na een halfwaardetijd is rond de 50 procent van het plateau bereikt, na twee rond de 75 procent, na drie rond de 87,5 procent en na vijf reeds meer dan 96 procent. Precies hetzelfde patroon dat de afbraak van een enkele toediening beschrijft, stuurt spiegelbeeldig ook de opbouw naar de steady state, waardoor beide processen exact even lang duren.
Doorslaggevend is een vaak verkeerd begrepen eigenschap: de tijd tot de steady state hangt uitsluitend af van de halfwaardetijd, niet van de doseringshoogte. Een hogere dosis leidt tot een hoger plateau, maar niet tot een sneller bereiken van het evenwicht. Bij tirzepatide met rond de vijf dagen halfwaardetijd wordt de steady state daarom pas na ongeveer vier weken wekelijkse toediening bereikt (Schneck et al., 2024). Bij peptiden met een zeer korte halfwaardetijd van enkele minuten daarentegen wordt na elke enkele toediening nagenoeg volledig geelimineerd, zodat zich nauwelijks een klassiek plateau opbouwt. Het concept van de steady state is dus alleen relevant voor stoffen waarvan de halfwaardetijd in het bereik van het doseringsinterval of daarboven ligt. Wie het plateau sneller wil bereiken zonder de onderhoudsdosis blijvend te verhogen, zou met een eenmalige oplaaddosis moeten werken, wat echter een afzonderlijk concept is buiten de zuivere halfwaardetijdbeschouwing.
De accumulatiefactor (Rac) kwantificeert met hoeveel de concentratie in de steady state toeneemt ten opzichte van een enkele toediening. De basisformule luidt Rac = 1 gedeeld door (1 min het in het interval resterende deel), of gelijkwaardig 1 gedeeld door het in het interval geelimineerde deel. Uitgedrukt met de halfwaardetijd ontstaat Rac = 1 gedeeld door (1 min 0,5 tot de macht (doseringsinterval gedeeld door t1/2)). Het in het interval resterende deel volgt op gelijkwaardige wijze uit e tot de macht min k maal tau, waarbij tau het doseringsinterval is.
Een concreet voorbeeld maakt dit duidelijk: bedraagt het doseringsinterval precies een halfwaardetijd, dan resteert aan het einde van elk interval 50 procent. De accumulatiefactor is dan 1 gedeeld door (1 min 0,5), gelijk aan 2, de steady-state-concentratie ligt dus ongeveer op het dubbele van een enkele toediening. Is het interval twee keer zo lang als de halfwaardetijd, dan resteert 25 procent en bedraagt Rac rond de 1,33. Bij zeer korte intervallen ten opzichte van de halfwaardetijd stijgt de factor sterk; bij een interval van een kwart van de halfwaardetijd bijvoorbeeld tot rond de 6,3. Echte gegevens passen bij dit model: tirzepatide vertoonde bij wekelijkse toediening een gemiddelde accumulatie van ongeveer 1,7-voudig, wat strookt met de halfwaardetijd van rond de vijf dagen en een interval van zeven dagen (Schneck et al., 2024). Hoe langer de halfwaardetijd in verhouding tot het interval, des te sterker de accumulatie.
In de farmacokinetische praktijk is de halfwaardetijd het belangrijkste aanknopingspunt voor de keuze van de afstand tussen herhaalde toedieningen in een onderzoeksprotocol. Een interval dat duidelijk korter is dan de halfwaardetijd leidt tot sterke accumulatie en hoge plateauconcentraties. Een interval dat een veelvoud van de halfwaardetijd bedraagt, laat de concentratie tussen de toedieningen ver dalen en veroorzaakt grote schommelingen tussen piek- en dalwaarden.
De reden voor dit verband is zuiver wiskundig: de verhouding tussen piek- en dalconcentratie in de steady state wordt uitsluitend bepaald door de verhouding tussen doseringsinterval en halfwaardetijd. Ligt het interval op een halfwaardetijd, dan halveert de concentratie tussen twee toedieningen, de verhouding tussen piek en dal bedraagt dus ongeveer 2 op 1. Bij een interval van vier halfwaardetijden daalt de concentratie tot een zestiende, de schommelingsbreedte wordt dus extreem. Juist de lange halfwaardetijden van moderne peptiden verklaren daarom hun doseringsschema's. Semaglutide met rond de een week halfwaardetijd (Yang et al., 2024) en tirzepatide met rond de vijf dagen (Schneck et al., 2024) laten intervallen in het weekbereik toe, omdat de concentratie tussen twee toedieningen niet te sterk daalt. Kortlevende peptiden zoals BPC-157 met een halfwaardetijd in minuten (He et al., 2022) zouden bij hetzelfde schema tussen de toedieningen praktisch volledig verdwijnen. De halfwaardetijd stelt dus het kader vast waarbinnen een zinvolle verhouding van schommelingsbreedte en accumulatie mogelijk is. Wie met verschillende peptiden werkt, kan via de peptide-calculator hoeveelheden en via de halfwaardetijd-calculator de tijdsverlopen modelleren.
Een volledig uitgewerkt voorbeeld verbindt de voorgaande concepten. Stel dat een onderzoekspeptide een halfwaardetijd van 48 uur heeft en elke 24 uur in dezelfde hoeveelheid wordt toegevoegd. Allereerst de eliminatiesnelheidsconstante: k = 0,693 gedeeld door 48, gelijk aan rond de 0,0144 per uur. Het in het interval van 24 uur resterende deel is e tot de macht min 0,0144 maal 24, dus rond de 0,707, waarmee ongeveer 70,7 procent resteert.
Daaruit volgt de accumulatiefactor: Rac = 1 gedeeld door (1 min 0,707), gelijk aan rond de 3,4. De steady-state-concentratie ligt dus ongeveer op het 3,4-voudige van wat een enkele toediening voortbrengt. Tot deze steady state is bereikt, verstrijken rond de vijf halfwaardetijden, dus ongeveer 240 uur of tien dagen. Verlengt men het interval tot 48 uur, dus precies een halfwaardetijd, dan daalt het resterende deel tot 50 procent en valt Rac terug op 2. Verkort men het daarentegen tot 12 uur, dus een kwart van de halfwaardetijd, dan stijgt het resterende deel tot rond de 0,841 en klimt Rac tot ongeveer 6,3. Dit cijferspel toont aanschouwelijk hoe gevoelig de accumulatie reageert op de verhouding tussen interval en halfwaardetijd. Precies deze berekeningen neemt de halfwaardetijd-calculator u uit handen en geeft kromme, steady state en accumulatiefactor grafisch weer. Een basisintroductie in de molecuulklasse biedt bovendien het artikel Wat zijn peptiden?.
Alle voorgaande berekeningen berusten op het een-compartimentmodel, dat het lichaam behandelt als een enkele, homogeen gemengde ruimte waaruit de stof met een enkele snelheidsconstante verdwijnt. Dit model is elegant omdat het met een enkele halfwaardetijd toekomt, en het is voor veel onderzoeksdoeleinden voldoende nauwkeurig. Het beeldt de werkelijkheid echter slechts bij benadering af, want het neemt aan dat een peptide zich onmiddellijk na de toediening direct en gelijkmatig over het gehele verdelingsvolume verdeelt.
In werkelijkheid verdelen veel peptiden zich niet ogenblikkelijk. Ze stromen eerst snel naar goed doorbloede weefsels en pas langzaam naar slechter doorbloede compartimenten. Daaruit ontstaat een tweefasig verloop: een steile verdelingsfase kort na de toediening, gevolgd door een vlakkere eliminatiefase waarin de concentratie wordt gehandhaafd door herverdeling vanuit de weefsels. Dergelijk gedrag beschrijft het twee-compartimentmodel nauwkeuriger en levert dan twee verschillende halfwaardetijden op, een korte verdelingshalfwaardetijd en een langere terminale halfwaardetijd. Wie slechts naar een enkel getal kijkt, verwart deze twee fasen gemakkelijk en onderschat de verblijfsduur, omdat de terminale fase de verblijfsduur in het weefsel bepaalt, terwijl de verdelingsfase slechts de aanvankelijke, snelle daling van de plasmaconcentratie weerspiegelt.
Juist hier ligt de praktische consequentie voor de modellering: de vuistregel van de vier tot vijf halfwaardetijden heeft betrekking op de terminale halfwaardetijd, niet op de snelle verdelingsfase. Wie ten onrechte de verdelingshalfwaardetijd inzet, zou de eliminatieduur dramatisch onderschatten. Verdere grenzen betreffen niet-lineaire kinetiek bij verzadiging van de eliminerende systemen, actieve metabolieten die zelf werkzaam zijn en de effectieve werkingsduur verlengen, alsook uitgesproken weefselbinding die de terminale fase verlengt. Ook de in de rekenvoorbeelden gebruikte aanname van een onmiddellijke, volledige resorptie geldt alleen voor de intraveneuze toediening; bij subcutane toediening vertraagt de resorptie het verloop bovendien en kan zij de schijnbare halfwaardetijd verlengen, een effect dat bekendstaat als flip-flopkinetiek. Voor de meeste onderzoekspeptiden in het gebruikelijke dosisbereik blijft het een-compartimentmodel niettemin een bruikbare benadering, maar men moet zich bewust zijn van de vereenvoudiging en vuistregels niet overbelasten.
Niet noodzakelijk. Een lange halfwaardetijd vlakt weliswaar concentratieschommelingen af en maakt minder frequente toedieningen mogelijk, maar verlengt ook de tijd tot de steady state en de verblijfsduur na het staken. Welke eigenschap wenselijk is, hangt volledig af van het onderzoeksdoel en laat zich niet in algemene termen beantwoorden.
Bij een eersteordekinetiek is de halfwaardetijd concentratieonafhankelijk en blijft zij over het gebruikelijke dosisbereik constant. Pas bij verzadiging van de eliminerende systemen, dus bij de overgang naar de nulde orde, kan de schijnbare halfwaardetijd dosisafhankelijk worden. Voor de meeste onderzoekspeptiden is dat niet relevant.
Gerichte moleculaire modificaties zoals de vervanging van aminozuren of het aanhechten van vetzuurketens verhogen de albuminebinding en beschermen tegen enzymatische afbraak (Menacho-Melgar et al., 2018). Semaglutide draagt bijvoorbeeld een C18-vetzuurdizuur-zijketen die de halfwaardetijd tot rond de 160 uur verlengt (Yang et al., 2024).
Het is een goede benadering voor stoffen met eersteordekinetiek, maar heeft uitzonderingen (Wadhwa & Cascella, 2023). Stoffen met niet-lineaire kinetiek, uitgesproken weefselbinding of actieve metabolieten kunnen duidelijk afwijken. De regel vervangt daarom geen stofspecifieke modellering.
Uitsluitend voor onderzoeksdoeleinden. Niet bestemd voor menselijke consumptie. Wetenschappelijke redactie: Dr. Sieglinde Klaus
